Virus inhibidor del tomàquet
El virus inhibidor del tomàquet (en anglès, Tomato Bushhy Stunt, TBSV) és un virus de la família dels Tombusviridae.[1]
| Tomato bushy stunt virus | |
|---|---|
 | |
| Dades | |
| Hoste | |
| Genoma | virus d'ARN monocatenari positiu, virus d'ARN monocatenari positiu i virus d'ARN monocatenari positiu |
| Taxonomia | |
| Família | Tombusviridae |
| Gènere | Tombusvirus |
| Espècie | Tomato bushy stunt virus |
Es va informar per primera vegada als tomàquets l'any 1935 i afecta principalment els cultius d'hortalisses, tot i que generalment no es considera un patogen vegetal econòmicament significatiu. Depenent de l'hoste, el TBSV provoca retard del creixement, taques de fulles i fruita deformada o absent. És probable que el virus es transmeti al sòl a l'entorn natural, però també es pot transmetre mecànicament, per exemple, mitjançant eines de tall contaminades.
El TBSV s'ha utilitzat com a sistema model en la investigació virològica sobre el cicle de vida dels virus vegetals, particularment en infeccions experimentals de la planta hoste model Nicotiana benthamiana.[2][3]
Gamma d'hostes
modificaEl TBSV té una àmplia gamma d'hostes en condicions experimentals i s'ha informat que infecta més de 120 espècies de plantes que abasten 20 famílies. No obstant això, en condicions naturals la seva gamma és molt més estreta i, en general, inclou hortalisses de cultiu i plantes ornamentals. Es va identificar per primera vegada a les plantes de tomàquet i també s'ha documentat que afecta la poma, la carxofa, la cirera, la vinya, el llúpol i el pebrot. Tot i que provoca una pèrdua important de rendiment a les plantes de tomàquet, no es considera un fitopatogen econòmicament significatiu.[4][3]
-
 N. benthamiana, un host experimental comú per a TBSV
N. benthamiana, un host experimental comú per a TBSV

Es tracta, però, d'un sistema model molt ben establert per a l'estudi de virus vegetals, generalment mitjançant la infecció experimental de Nicotiana benthamiana o Nicotiana clevelandii, parents de plantes de tabac en les quals el TBSV pot causar infecció sistèmica. En particular, la planta model comú Arabidopsis thaliana no és un hoste.[2][3] El TBSV també es pot replicar en rents en condicions de laboratori.[5]
Símptomes
modificaEls signes de TBSV depenen de l'hoste. Les infeccions locals poden provocar lesions necròtiques o cloròtiques. Les infeccions sistèmiques poden provocar un creixement retardat, fruits deformats o absents i fulles danyades; en entorns agrícoles el rendiment es pot reduir significativament. L'aparença raquítica i «tupida» de les plantes de tomàquet en què es va descobrir el virus va donar el nom al patogen. En alguns hostes, sobretot N. benthamiana, el TBSV pot causar necrosi sistèmica letal.[4][3]
Transmissió
modificaEs creu que el TBSV es transmet de manera passiva a la natura, principalment a través del sòl o l'aigua. No hi ha organismes vectors coneguts; la transmissió per pugons, àcars i el fong Olpidium brassicae s'ha descartat específicament.[4]
Tanmateix, s'ha observat que el virus de la necrosi del cogombre (Cucumber necrosis virus, CNV) molt relacionat amb el tombusvirus es transmet per zoòspores d'Olpidium bornovanus, de manera que la transmissió del TBSV per un vector encara desconegut segueix sent una possibilitat.[3]
El TBSV també es pot transmetre per llavor o per inoculació mecànica.[4][3] En proves experimentals, el virus pot sobreviure al pas pel sistema digestiu humà si es consumeix en els aliments i seguirà sent infecciós; s'ha plantejat la hipòtesi que es podria produir la propagació per aigües residuals.[6]
Distribució i gestió
modificaEl TBSV es distribueix bastant àmpliament per l'Europa central i occidental, el nord d'Àfrica i Amèrica del Nord i del Sud.[4][3] No es recomana cap mesura específica de control per al virus, tot i que les directrius de control de plagues distribuïdes per la Universitat de Califòrnia recomanen evitar camps amb antecedents de TBSV o utilitzar rotacions llargues de cultius.[7]
Taxonomia
modificaEl TBSV s'assigna al gènere Tombusvirus de la família Tombusviridae.[8] Tant el gènere com la família deriven els seus noms d'una abreviatura del seu nom en anglès, «tomato bushy stunt virus».[9]
Estructura
modificaEl TBSV és un virus icosaèdric sense embolcall amb una càpsida vírica T=3 composta per 180 subunitats d'una sola proteïna de la càpsida. La seva estructura va ser estudiada àmpliament per cristal·lografia de raigs X a partir de finals de la dècada del 1950; la seva simetria icosaèdrica la va identificar per primera vegada el biòleg estructural Donald Caspar, que també va ser pioner en l'estudi del virus del mosaic del tabac.[10]
-
 Dibuixos esquemàtics d'un virió Tombusviridae (secció transversal i vista lateral)
Dibuixos esquemàtics d'un virió Tombusviridae (secció transversal i vista lateral)

Un mapa de resolució gairebé atòmica va ser obtingut el 1978 per un equip d'investigació que va incloure Stephen C. Harrison.[11][12]
Complement del genoma i proteïna
modificaEl TBSV és un virus d'ARN monocatenari de sentit positiu amb un genoma lineal de ~ 4800 nucleòtids.[13][14]
El genoma conté cinc gens que codifiquen una replicasa composta per dues proteïnes (p33 i p92), una proteïna de la càpsida (anomenada CP o p41) i dues proteïnes addicionals, el supressor de silenciament de l'ARN p19 i la proteïna de moviment p22.[3]
-
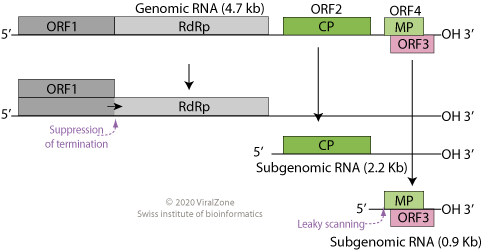 Mapa del genoma del Tombusvirus
Mapa del genoma del Tombusvirus

Aquestes dues proteïnes s'expressen a partir de gens superposats disposats de manera que el marc obert de lectura de p19 estigui completament dins de l'ORF de p22.[15] El genoma conté un possible gen addicional, anomenat pX, de funció desconeguda.[3]
p33 i p92 (replicasa)
modificaJunts, p33 i p92 formen el complex de replicasa viral. P33 és més petit i p92 es produeix a través de la lectura ribosòmica del codó d'aturada p33, donant lloc a una seqüència d'aminoàcids N-terminal compartida i un gran excés de p33 en relació amb p92. Les proteïnes p33 s'uneixen de manera cooperativa als àcids nucleics monocatenaris, mentre que la proteïna p92 és una ARN polimerasa depenent de l'ARN (RdRp). Tots dos són essencials per a la proliferació viral. Ambdues proteïnes estan associades a les membranes cel·lulars.[3]
p41 (proteïna de la càpsida)
modificaLa proteïna de la càpsida viral CP, o p41, és una doble proteïna de gelatina que s'assembla en una càpsida icosaèdrica que conté 180 còpies de la proteïna. La formació de virions no sempre és necessària per a la propagació localitzada del virus a les cèl·lules vegetals veïnes, perquè les partícules de ribonucleoproteïna que contenen material genètic viral es poden estendre als veïns immediats a través dels plasmodesmes. Tanmateix, la proteïna de la càpsida és necessària per a la infecció sistèmica.[3]
p19 (supressor de silenciament de l'ARN)
modificaLa proteïna p19 és un factor de patogenicitat i funciona suprimint la via de silenciament de l'ARN, una forma habitual de defensa antiviral. La proteïna p19 s'uneix als ARN interferents petits i impedeix la seva incorporació al complex de silenciament induït per l'ARN (RNA-induced silencing complex, RISC), permetent així la propagació viral a la planta hoste.[2][16][17]
-
 Complex de la proteïna p19 amb ARN de doble cadena
Complex de la proteïna p19 amb ARN de doble cadena

La presència de p19 és necessària per a la infecció sistèmica o per a la infecció letal en alguns hostes; a l'hoste experimental N. benthamiana, p19 media en gran manera la necrosi sistèmica letal que és el resultat de la infecció per TBSV.[3][16]
p22 (proteïna de moviment)
modificaLa proteïna p22 és una proteïna de moviment necessària perquè el virus es propagui de cèl·lula a cèl·lula. P22 és una proteïna d'unió a l'ARN que s'associa amb la paret cel·lular i facilita el moviment del material genètic viral d'una cèl·lula a la seva veïna mitjançant plasmodesmes interconnectats.[3]
Replicació
modificaUn virió TBSV conté una còpia del seu genoma d'ARN monocatenari de sentit positiu, que és lineal i no té una cua de poliadenina de 3' o una caputxa 5'. No obstant això, les proteïnes p33 i p92 es tradueixen directament a partir de l'ARN genòmic. Quan el genoma es replica, es produeixen dues molècules d'ARN subgenòmic que actuen com a ARN missatger; un a partir del qual s'expressa el gen p41 (CP) i un dels gens p19 i p22. Els gens p19 i p22 superposats es tradueixen ambdós a través dels efectes de l'escaneig amb fuites.[3] S'han identificat diverses interaccions a llarga distància entre àrees linealment ben separades del genoma amb importància funcional per garantir una replicació eficient.[15]
ARN interferent defectuós
modificaLes molècules d'ARN interferent defectuoses (DI) són ARN que es produeixen a partir del genoma viral però que no són competents per infectar cèl·lules per si soles; en canvi, requereixen coinfecció amb un virus «auxiliar» intacte. Les infeccions per TBSV sovint produeixen un nombre significatiu de DIs de parts consistents del genoma en condicions experimentals, però aquest comportament no s'ha observat a la natura. És probable que la seva producció sigui específica de l'amfitrió. Les infeccions que donen lloc a DIs solen tenir signes més lleus.[3][18]
Referències
modifica- ↑ Mahy, van Regenmortel i 2009, p0445 i seg..
- ↑ 2,0 2,1 2,2 Scholthof, 2006, p. 405-411.
- ↑ 3,00 3,01 3,02 3,03 3,04 3,05 3,06 3,07 3,08 3,09 3,10 3,11 3,12 3,13 3,14 Yamamura i Scholthof, 2005, p. 491-502.
- ↑ 4,0 4,1 4,2 4,3 4,4 Martelli, G. P.; Russo, M.; Rubino, L. «Tomato bushy stunt virus» (en anglès). Descriptions of Plant Viruses (Association of Applied Biologists), desembre 2001.
- ↑ Balique et al., 2015, p. 2074-2098.
- ↑ «Integrated Pest Management Program» (en anglès). UC Pest Management Guidelines (University of California. Division of Agriculture and Natural Resources), desembre 2013.
- ↑ «ICTV. Taxonomy History for Tomato bushy stunt virus» (en anglès). International Committee on Taxonomy of Viruses, juliol 2015.
- ↑ Harrison et al., Shepherd, p. 356-363.
- ↑ Rossmann, 2013, p. 133-180.
- ↑ Harrison et al., Bricogne, p. 368-373.
- ↑ Hellemans i Bunch, 1988.
- ↑ Hearne et al., 1990, p. 141-151.
- ↑ Wagner et al., 2007.
- ↑ 15,0 15,1 Wu et al., White, p. e1003363.
- ↑ 16,0 16,1 Scholthof et al., 1995, p. 425-438.
- ↑ Jones, Jackson i Morris, 1990, p. 539-545.
- ↑ Scholthof, Scholthof i Jackson, 1995, p. 324-328.
Bibliografia
modifica- Balique, Fanny; Lecoq, Hervé; Raoult, Didier; Colson, Philippe «Can Plant Viruses Cross the Kingdom Border and Be Pathogenic to Humans?» (en anglès). Viruses, 7(4), abril 2015. DOI: 10.3390/v7042074. PMC: 4411691. PMID: 25903834.
- Harrison, B. D.; Finch, J. T.; Gibbs, A. J.; Hollings, M.; Shepherd, R. J.; Valenta, V.; Wetter, C. «Sixteen groups of plant viruses» (en anglès). Virology, 45(2), agost 1971. DOI: 10.1016/0042-6822(71)90336-9. PMID: 5106891.
- Harrison, S. C.; Olson, A. J.; Schutt, C. E.; Winkler, F. K.; Bricogne, G. «Tomato bushy stunt virus at 2.9 Å resolution» (en anglès). Nature, 276(5686), novembre 1978. Bibcode: 1978Natur.276..368H. DOI: 10.1038/276368a0. PMID: 19711552.
- Hearne, Patrick Q.; Knorr, David A.; Hillman, Bradley I.; Morris, Thomas J. «The complete genome structure and synthesis of infectious RNA from clones of tomato bushy stunt virus» (en anglès). Virology, 177(1), juliol 1990. DOI: 10.1016/0042-6822(90)90468-7. PMID: 2353450.
- Hellemans, Alexander; Bunch, Bryan H. The timetables of science: a chronology of the most important people and events in the history of science (en anglès). Simon & Schuster, 1988. ISBN 978-0-671-62130-8.
- Jones, Richard W.; Jackson, A.O.; Morris, Thomas J. «Defective-interfering RNAs and elevated temperatures inhibit replication of tomato bushy stunt virus in inoculated protoplasts» (en anglès). Virology, 176(2), 1990. DOI: 10.1016/0042-6822(90)90024-L. PMID: 2345965.
- Mahy, Brian W. J.; van Regenmortel, Marc H. V.. Desk Encyclopedia of Plant and Fungal Virology (en anglès). Academic Press, 2009. ISBN 978-0-12-375148-5.
- Rossmann, Michael G. «Structure of viruses: a short history» (en anglès). Quarterly Reviews of Biophysics, 46(2), maig 2013. DOI: 10.1017/S0033583513000012. ISSN: 0033-5835. PMID: 23889891.
- Scholthof, Karen-Beth G.; Scholthof, Herman B.; Jackson, Andrew O. «The Effect of Defective Interfering RNAs on the Accumulation of Tomato Bushy Stunt Virus Proteins and Implications for Disease Attenuation» (en anglès). Virology, 211(1), agost 1995. DOI: 10.1006/viro.1995.1410. PMID: 7645230.
- Scholthof, Herman B.; Scholthof, Karen-Beth G.; Kikkert, Marjolein; Jackson, A. O. «Tomato Bushy Stunt Virus Spread Is Regulated by Two Nested Genes That Function in Cell-to-Cell Movement and Host-Dependent Systemic Invasion» (en anglès). Virology, 213(2), 1995. DOI: 10.1006/viro.1995.0015. PMID: 7491767.
- Scholthof, Herman B. «The Tombusvirus-encoded P19: from irrelevance to elegance» (en anglès). Nature Reviews Microbiology, 4(5), març 2006. DOI: 10.1038/nrmicro1395. PMID: 16518419.
- Wagner, Edward K.; Hewlett, Martinez J.; Bloom, David C.; Camerini, David. Basic Virology (en anglès). John Wiley & Sons, 2007. ISBN 9781405147156.
- Wu, Baodong; Grigull, Jörg; Ore, Moriam O.; Morin, Sylvie; White, K. Andrew «Global Organization of a Positive-strand RNA Virus Genome» (en anglès). PLOS Pathogens, 9(5), maig 2013. DOI: 10.1371/journal.ppat.1003363. ISSN: 1553-7374. PMC: 3662671. PMID: 23717202.
- Yamamura, Y.; Scholthof, H. B. «Tomato bushy stunt virus: a resilient model system to study virus-plant interactions» (en anglès). Molecular Plant Pathology, 6(5), setembre 2005. DOI: 10.1111/j.1364-3703.2005.00301.x. PMID: 20565674.