Regions HNLC
Les regions altament nutritives i baixes de clorofil·la (en anglès, High-nutrient, low-chlorophyll regions, o HNLC regions) són regions de l'oceà on l'abundància de fitoplàncton és baixa i bastant constant malgrat la disponibilitat de macronutrients. El fitoplàncton depèn d'un conjunt de nutrients per a la funció cel·lular. Els macronutrients (per exemple, nitrats, fosfats, àcids silícics) estan generalment disponibles en quantitats superiors en aigües superficials de l'oceà i són els components típics dels adobs comuns del jardí. Els micronutrients (per exemple, ferro, zinc, cobalt) estan generalment disponibles en quantitats menors i inclouen oligoelements. Els macronutrients solen estar disponibles en concentracions milimolars, mentre que els micronutrients estan generalment disponibles en concentracions de micro i nanomolar. En general, el nitrogen tendeix a ser un nutrient oceànic limitant, però en les regions HNLC mai no està esgotat significativament.[1][2] En canvi, aquestes regions solen estar limitades per baixes concentracions de ferro metabolitzable.[1] El ferro és un micronutrient crític del fitoplàncton, necessari per a la catàlisi enzimàtica i el transport d'electrons.[3][4]
Entre la dècada del 1930 i la dècada del 1980, es va plantejar la hipòtesi que el ferro és un micronutrient oceànic limitant, però no hi havia mètodes suficients per detectar de manera fiable el ferro a l'aigua de mar per confirmar aquesta hipòtesi.[5] El 1989 es van detectar altes concentracions de sediments rics en ferro a les aigües costaneres properes al golf d'Alaska.[6] No obstant això, les aigües de la costa tenien baixes concentracions de ferro i una menor productivitat malgrat la disponibilitat de macronutrients per al creixement del fitoplàncton.[6] Aquest patró es va observar en altres regions oceàniques i va donar lloc a la designació de tres zones HNLC principals: Oceà Pacífic nord, Oceà Pacífic equatorial i Oceà Antàrtic.[1][2]
El descobriment de les regions HNLC ha fomentat el debat científic sobre l'ètica i l'eficàcia dels experiments de fertilització amb ferro que intenten reduir el diòxid de carboni atmosfèric estimulant la fotosíntesi a nivell de superfície. També ha donat lloc al desenvolupament d'hipòtesis com el control del pasturatge que suposa que les regions HNLC es formen, en part, del pasturatge del fitoplàncton (per exemple, dinoflagel·lats, ciliats) per organismes més petits (per exemple, protists).
Producció primària
modifica
La producció primària és el procés pel qual els autòtrofs utilitzen la llum per convertir el carboni del diòxid de carboni aquós en sucre per al creixement cel·lular.[7] La llum catalitza el procés fotosintètic i els nutrients s'incorporen a la matèria orgànica. Perquè es produeixi la fotosíntesi, els macronutrients com el nitrat i el fosfat han d'estar disponibles en proporcions suficients i en formes biodisponibles per a la seva utilització. La relació molecular de 106 (carboni): 16 (nitrogen): 1 (fòsfor) va ser descoberta per Redfield, Ketcham i Richards (RKR) i es coneix com la relació de Redfield.[8]
La fotosíntesi ( ) i la respiració ( ) es representa amb l'equació:[9]



La fotosíntesi pot estar limitada per deficiències de certs macronutrients. No obstant això, al Pacífic Nord, Pacífic Equatorial i Oceà Antàrtic, els macronutrients es troben en proporcions, quantitats i formes biodisponibles suficients per donar suport a nivells més elevats de producció primària del que es va trobar. La disponibilitat de macronutrients a les regions HNLC juntament amb la biomassa baixa de fitoplàncton suggereix que algun altre procés biogeoquímic limita el creixement del fitoplàncton.[7]
Atès que la producció primària i la biomassa del fitoplàncton no es poden mesurar actualment sobre conques oceàniques senceres, els científics utilitzen la clorofil·la α com a representant de la producció primària. Les observacions modernes dels satèl·lits controlen i rastregen les abundàncies globals de la clorofil·la α a l'oceà mitjançant la teledetecció. Les concentracions més altes de clorofil·la, generalment indiquen àrees de producció primària alta i, per contra, els nivells inferiors de clorofil·la indiquen una producció primària baixa. Aquesta coincidència de baixa clorofil·la i una alta disponibilitat de macronutrients és la raó per la qual es considera que aquestes regions són «altament nutritives i baixes de clorofil·la».
A més dels macronutrients necessaris per a la síntesi de matèria orgànica, el fitoplàncton necessita micronutrients, com ara oligoelements per a funcions cel·lulars.[7] La disponibilitat de micronutrients pot restringir la producció primària perquè els restes de metalls de vegades limiten els nutrients. S'ha determinat que el ferro és un micronutrient limitant principal en les regions HNLC.[5] Estudis recents han indicat que el zinc i el cobalt poden ser micronutrients secundaris i / o co-limitants.[10][11]
Distribució global
modificaCaracterístiques comunes
modificaLes regions HNLC cobreixen el 20% dels oceans del món i es caracteritzen per tenir diferents patrons físics, químics i biològics. Aquestes aigües superficials tenen concentracions de macronutrients variables, però relativament abundants anualment, comparades amb altres regions oceàniques.[5] Mentre que les regions HNLC descriuen àmpliament les tendències biogeoquímiques d'aquestes grans regions oceàniques, les tres zones experimenten floracions estacionals de fitoplàncton en resposta als patrons atmosfèrics globals. De mitjana, les regions HNLC solen tenir limitat el creixement del ferro i el zinc és variable.[11][12] Aquesta limitació d'oligoelements de metalls produeixen comunitats de fitoplàncton de menor grandària. En comparació amb les regions més productives de l'oceà, les zones HNLC tenen proporcions més altes d'àcid silícic respecte al nitrat, ja que les diatomees més grans, que requereixen àcid silícic per fer les seves closques d'òpal (SiO₂·nH₂O), són menys freqüents.[10][11][12] A diferència de l'Oceà Pacífic sud i del Pacífic nord, el Pacífic equatorial experimenta una disponibilitat temporal de silicats que condueix a grans floracions estacionals de diatomees.[13][14]
-
 Concentració mitjana d'àcid silícic superficial
Concentració mitjana d'àcid silícic superficial
(mmol Si m-3) -
 Concentració mitjana de nitrat superficial
Concentració mitjana de nitrat superficial
(mmol N m-3) -
 Concentració mitjana de fosfat superficial
Concentració mitjana de fosfat superficial
(mmol P m-3)
.png)
.png)
.png)
La distribució d'oligoelements i l'abundància relativa de macronutrients es reflecteixen en l'estructura de la comunitat de plàncton. Per exemple, la selecció del fitoplàncton amb una elevada relació superfície/volum fa que les regions HNLC estiguin dominades per nanoplàncton i picoplàncton. Aquesta relació permet una utilització òptima dels nutrients dissolts disponibles. El fitoplàncton més gran, com les diatomees, no es pot mantenir enèrgicament en aquestes regions. El picoplàncton comú dins d'aquestes regions inclouen gèneres com el Prochlorococcus (que no es troba generalment al Pacífic nord), el synechococcus i diverses eucariotes. Els protists de pasturatge probablement controlen l'abundància i distribució d'aquests petits fitoplàncton.[15][16]
La producció primària neta generalment més baixa a les zones HNLC provoca una disminució de la reducció biològica del diòxid de carboni atmosfèric i, per tant, aquestes regions són generalment considerades una font neta de diòxid de carboni atmosfèric.[14] Les àrees HNLC són d'interès per als geoenginyers i per alguns de la comunitat científica que creuen que la fertilització de grans superfícies d'aquestes aigües amb ferro podria reduir el diòxid de carboni dissolt i compensar les emissions de carboni antropogèniques.[6] L'anàlisi de les dades del nuclis de gel antàrtic mostren una correlació durant els últims milions d'anys entre els alts nivells de pols i la baixa temperatura, indicant que l'addició de pols rics en ferro al mar ha estat un amplificador natural del refredament del clima.[17]
El Pacífic nord
modificaEl descobriment i el nomenament de la primera regió HNLC, el Pacífic nord, es va formalitzar en un article fonamental publicat el 1988.[6] L'estudi va concloure que les aigües superficials del Pacífic nord oriental són generalment dominades pel picoplàncton, tot i la relativa abundància de macronutrients.[6] En altres paraules, no es va trobar fitoplàncton més gran, com les diatomees que es desenvolupen en aigües riques en nutrients. En canvi, les aigües superficials estaven repletes de picoplàncton i nanoplàncton més petits.[6] Basant-se en experiments de nutrients de laboratori, es va pensar que el ferro era un micronutrient limitant clau.[6]
L'oceà Pacífic és el cos d'aigua més gran i més antic de la Terra. El Pacífic nord es caracteritza per la rotació generalitzada en sentit horari del gir del Pacífic nord, que és impulsada pels vents alisis. Les variacions espacials en els vents alisis produeixen temperatures de l'aire més fresques al Pacífic nord-oest i temperatures de l'aire més suaus al Pacífic nord-oriental (és a dir, al Pacífic subàrtic).[18] El ferro és subministrat al Pacífic nord per les tempestes de pols que es produeixen a Àsia i Alaska, així com aigües riques en ferro adveccionades des del marge continental, de vegades per remolins com els Haida Eddies.[19][20]
-

-
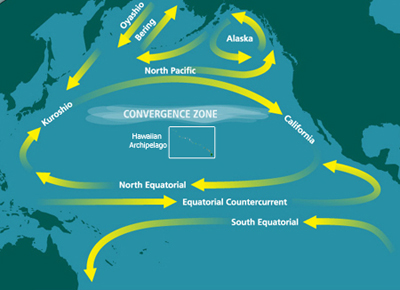 Corrents al Pacífic Nord.
Corrents al Pacífic Nord.
.jpg)

No obstant això, les concentracions de ferro varien al llarg de l'any. Els corrents oceànics són impulsats per patrons atmosfèrics estacionals que transporten el ferro des del marge de la fossa de les Kurils-Kamtxatka cap al Pacífic subàrctic occidental. Aquesta introducció del ferro proporciona un subministrament per sota de la superfície de micronutrients, que poden ser utilitzats pels productors primaris durant la remodelació de les aigües més profundes a la superfície.[21] La profunditat del fons marí també pot estimular la floració del fitoplàncton a les regions HNLC, ja que el ferro es difon del fons marí i alleuja la limitació de ferro en aigües poc profundes.[22] Les investigacions dutes a terme al golf d'Alaska van mostrar que les zones amb aigües poc profundes, com la plataforma sud d'Alaska, tenen una intensitat de fitoplàncton més intensa que les aigües profundes.[22] Les cendres volcàniques procedents de l'erupció del volcà Kasatochi a l'agost de 2008 van proporcionar un exemple de fertilització natural amb ferro a l'Oceà Pacífic nord-est.[23] La regió va ser fecundada per la pluja de pols volcànica que contenia ferro soluble. En els dies següents, les floracions de fitoplàncton eren visibles des de l'espai.[23]
Les limitacions de les concentracions d'oligoelements al Pacífic nord limiten les floracions de diatomees durant tot l'any.[24] Tot i que el Pacífic nord és una regió HNLC, produeix i exporta a l'interior de l'oceà una quantitat relativament elevada de sílice biogènic en partícules en comparació amb l'Atlàntic nord, que suporta un creixement significatiu de diatomees.[24]
El Pacífic equatorial
modificaEl Pacífic equatorial és una regió oceànica caracteritzada per un aflorament de gairebé tot l'any, a causa de la convergència dels vents alisis del nord-est i del sud-est a la zona de convergència intertropical. El Pacífic equatorial abasta gairebé la meitat de la circumferència de la Terra i té un paper important en la nova producció primària marítima mundial.[25] La «nova producció» és un terme utilitzat en l'oceanografia biològica per descriure la manera en què el nitrogen es recicla a l'oceà.[18] A les regions de nova producció millorada, els nitrats de la zona afòtica es converteixen en aigües superficials, reemplaçant el subministrament de nitrats de la superfície. Malgrat la disponibilitat de nitrogen a les aigües del Pacífic equatorial, la producció primària i la biomassa oceànica superficial observada és considerablement menor en comparació amb altres regions principals de l'oceà.[26] Així, el Pacífic equatorial és considerat com una de les tres principals regions HNLC.
Igual que altres grans regions HNLC, el Pacífic equatorial es considera limitat pels nutrients a causa de la manca d'oligoelements com el ferro. El Pacífic equatorial rep aproximadament entre 7 i 10 vegades més ferro de la sotacorrent equatorial (EUC) que de les entrades degudes a la solució de la pols atmosfèrica.[27] Les reconstruccions climàtiques dels períodes glacials mitjançant registres de proxy de sediments han revelat que el Pacífic equatorial podria haver estat 2,5 vegades més productiu que el modern oceà equatorial.[27] Durant aquests períodes glacials, el Pacífic equatorial va augmentar la seva exportació de nova producció marina,[27] proporcionant així un augment del diòxid de carboni atmosfèric. La ciència de la paleoceanografia intenta entendre la interacció dels cicles glacials amb la dinàmica oceànica. Els paleoceanògrafs actualment qüestionen la hipòtesi de la pols eòlica, que suggereix que el transport de pols atmosfèrica rica en ferro de l'Amèrica Central i Amèrica del Sud controla la intensitat de la producció primària al Pacífic equatorial.[27] Un estudi suggereix que, a causa que l'aflorament de l'EUC, proporciona la major part del ferro biodisponible a les aigües superficials equatorials; l'únic mètode per revertir les condicions HNLC és millorar l'aflorament.[28][27] En altres paraules, la millora de l'aflorament regional, en lloc de la deposició de pols atmosfèrica rica en ferro, pot explicar per què aquesta regió experimenta una productivitat primària més alta durant els períodes glacials.
En comparació amb el Pacífic nord i l'oceà Antàrtic, les aigües del Pacífic equatorial tenen nivells relativament baixos de sílice biogènica i, per tant, no admeten les existències significatives de diatomees.[14] El picoplàncton és el principal productor primari marí d'aquestes regions degut principalment a la seva capacitat d'assimilar les baixes concentracions d'oligoelements.[14] Diverses comunitats de fitoplàncton al Pacífic equatorial es pasturen a la mateixa velocitat que la seva taxa de creixement, la qual cosa limita encara més la producció primària.[28]
No existeix un consens actual sobre quina de les dues hipòtesis principals (pasturatge o micronutrients) controla la producció en aquestes aigües equatorials. És probable que les limitacions d'oligoelements seleccionin els organismes de petites cèl·lules, cosa que augmenta la pressió de pasturatge dels protists.[28] Mentre que el Pacífic equatorial manté característiques HNLC, la productivitat pot ser alta a vegades. La productivitat condueix a l'abundància d'aus marines. El Pacífic equatorial conté les zones pesqueres més grans del món de tonyina,[18] i és la llar del dofí tacat tropical.
L'oceà Antàrtic
modificaL'oceà Antàrtic és la regió HNLC més gran de l'oceà global. Les aigües superficials de l'oceà Antàrtic han estat àmpliament identificades com a riques en macronutrients, malgrat les poblacions de fitoplàncton baixes.[29][30][31] El ferro dipositat a l'Atlàntic nord s'incorpora a les aigües profundes de l'Atlàntic nord i es transporta a l'oceà Antàrtic a través de la circulació termohalina.[32] Finalment, es barreja amb l'aigua circumpolar antàrtica, el subministrament de l'aigua proporciona ferro i macronutrients a les aigües superficials de l'oceà Antàrtic. Per tant, les aportacions de ferro i la producció primària a l'oceà Antàrtic són sensibles a la pols sahariana rica en ferro que es diposita sobre l'Atlàntic. A causa de la baixa aportació de pols atmosfèrica directament a les aigües superficials de l'oceà Antàrtic,[33][34] les concentracions de clorofil·la α són baixes. La disponibilitat de llum a l'oceà Antàrtic canvia dràsticament estacionalment, però no sembla una restricció significativa en el creixement del fitoplàncton.[3]
-
 Un esquema dels corrents antàrtics.
Un esquema dels corrents antàrtics. -
 Circulació termohalina. Els camins blaus representen corrents d'aigua profunds, mentre que els camins vermells representen corrents superficials.
Circulació termohalina. Els camins blaus representen corrents d'aigua profunds, mentre que els camins vermells representen corrents superficials.


Els macronutrients presents a les aigües superficials de l'oceà Antàrtic provenen d'aigües profundes ascendents. Mentre que els micronutrients (com el zinc i el cobalt) poden limitar el creixement del fitoplàncton a l'oceà Antàrtic, el ferro sembla un micronutrient limitant crític.[4] Algunes regions de l'oceà Antàrtic experimenten concentracions adequades de ferro i macronutrients, però el creixement del fitoplàncton és limitat. Els estudis hidrogràfics[35][36] i les exploracions de la regió del Passatge del Sud de Drake[37] han observat aquest fenomen al voltant de les illes Crozet, les illes Kerguelen i les illes de Geòrgia del Sud i Sandwich del Sud.[38][37] Aquestes zones són adjacents a les regions de l'Antàrtida continental i de les illes de l'oceà Antàrtic. Es creu que els micronutrients necessaris per al creixement d'algues s'obtenen de la pròpia plataforma continental.[37] Llevat de zones properes a la plataforma antàrtica, la deficiència de micronutrients limita severament la productivitat a l'oceà Antàrtic.
La disponibilitat de ferro no és l'únic regulador de la productivitat del fitoplàncton i de la biomassa.[39][40] A l'oceà Antàrtic, es creu que les temperatures baixes predominants tenen un impacte negatiu sobre les taxes de creixement del fitoplàncton.[39] La taxa de creixement del fitoplàncton és molt intensa i de curta durada a les zones obertes envoltades de gel marí i de zones permanents de gel marí. Es creu que el pasturatge d'herbívors, com el krill, els copèpodes i els sàlpids, suprimeix l'estoc de fitoplàncton. A diferència de les aigües obertes de l'oceà Antàrticl, el pasturatge al llarg dels marges de la plataforma continental és baix, per la qual cosa la major part del fitoplàncton que no es consumeix s'enfonsa al fons del mar i proporciona nutrients als organismes bentònics.[40]
Hipòtesis
modificaTenint en compte la localització remota de les àrees HNLC, els científics han combinat dades de modelització i observació per estudiar els límits de la producció primària. La combinació d'aquestes dues fonts de dades permet comparar el Pacífic nord, el Pacífic equatorial i l'oceà Antàrtic. Les dues explicacions actuals per a les regions HNLC globals són les limitacions de creixement a causa de la disponibilitat de ferro i els controls de pasturatge del fitoplàncton.
Hipòtesi del ferro
modificaLa primera especulació que el fitoplàncton al «desert antàrtic» (part de l'oceà Antàrtic anomenat així a causa de la baixa biomassa del fitoplàncton) era limitat pel ferro va ser feta pel biòleg anglès Joseph Hart el 1930.[41] No obstant això, aquest debat només es va reprendre el 1980 per l'oceanògraf John Martin, quan va realitzar diverses anàlisis de nutrients sobre l'aigua de mar i va concloure que el ferro limitava el creixement del fitoplàncton en aquestes regions del «desert de l'Antàrtida». El 1988, John Martin va confirmar la hipòtesi que el ferro limita les floracions del fitoplàncton i les taxes de creixement al Pacífic nord. El seu treball es va extrapolar a altres regions HNLC mitjançant proves que relacionaven la baixa concentració de ferro a la superfície amb la baixa clorofil·la.[6]
A partir d'aquest moment, la comunitat oceanogràfica va començar a estudiar de forma més sistemàtica l'efecte d'aquestes grans àrees de l'oceà global sobre la productivitat primària i l'exportació de carboni cap a l'oceà profund. El 1988, John Gribbin va ser el primer científic a suggerir públicament que l'efecte d'hivernacle es podria reduir si s'afegeixen grans quantitats de compostos de ferro solubles en aigua com a fertilitzant del fitoplàncton. Però John Martin va ser més conegut després de la seva declaració: «Dona'm la meitat d'un petrolier carregat de ferro i et donaré una glaciació».[42] Aquesta frase es va donar en una conferència a l'Institut Oceanogràfic Woods Hole al juliol de 1988, i va marcar el començament de dècades d'experiments de fertilització oceànica. Tots aquests experiments van suposar que dispersar una quantitat relativament petita de ferro a les zones HNLC generaria grans floracions d'algues. Hipotèticament, amb un nombre suficient d'aquestes zones fertilitzades, el creixement de microalgues podria eliminar tant diòxid de carboni de l'atmosfera que l'efecte d'hivernacle es podria invertir i la superfície de la Terra es refredaria.
Entre 1993 i 2012 es van dur a terme una sèrie d'experiments de fertilització oceànica amb ferro, que van provocar de fitoplàncton, augmentant així l'exportació de carboni cap a l'oceà profund:[43][44][45]
|
Nom de l'experiment |
Oceà |
Any |
Quantitat de ferro afegit |
|
IronEx I (Iron experiment) |
Pacífic equatorial |
1993 |
450 kg |
|
IronEx II (Iron experiment) |
Pacífic equatorial |
1995 |
450 kg |
|
SOIREE (Southern Ocean iron release experiment) |
Antàrtic |
1999 |
1.750 kg |
|
EisenEx (Iron experiment) |
Antàrtic |
2000 |
2.350 kg |
|
SEEDS I (Subarctic Pacific iron experiment for ecosystem dynamics study) |
Pacífic nord |
2001 |
350 kg |
|
SOFeX (Southern Ocean iron experiment - north) |
Antàrtic |
2002 |
1.300 kg |
|
SOFeX (Southern Ocean iron experiment - south) |
Antàrtic |
2002 |
1.700 kg |
|
SERIES (Subarctic ecosystem response to iron enrichment study) |
Pacífic nord |
2002 |
490 kg |
|
SEEDS II (Subarctic Pacific iron experiment for ecosystem dynamics study) |
Pacífic nord |
2004 |
480 kg |
|
EIFEX (European iron fertilization experiment) |
Antàrtic |
2004 |
2.820 kg |
|
SAGE (Surface-ocean lower-atmosphere studies of air-sea gás exchange) |
Antàrtic |
2004 |
1.100 kg |
|
FeeP (Phosphate and iron addition experiment)* |
Atlàntic |
2004 |
1.840 kg |
|
CROZEX (Crozet natural iron bloom and export experiment) |
Antàrtic |
2005 |
|
|
LOHAFEX (Indian and German iron fertilization experiment) |
Antàrtic |
2009 |
20.000 kg |
|
HSRC (The Haida Salmon Restoration Corporation) |
Pacífic nord |
2012 |
120.000 kg |
En resposta a aquests experiments de fertilització amb ferro en àrees HNLC, es van observar grans respostes de fitoplàncton, com la disminució de la concentració de nutrients a la superfície i l'augment de l'activitat biològica.[46][47][48][49][50]
Els estudis realitzats han demostrat que fertilitzant amb ferro a intervals repetits durant la setmana van produir una resposta biològica més gran que realitzant només una gran fertilització.[47][49][51] La mida de la resposta biològica tendeix a dependre de les característiques biològiques, químiques i físiques d'un lloc. En el Pacífic nord i equatorial, es creu que la sílice restringeix la producció addicional després de la fertilització amb ferro, mentre que la llum limita la producció addicional a l'oceà Antàrtic.[47] El fitoplàncton més petit va ser el primer que va respondre a un augment del ferro, però van ser avançats ràpidament pel fitoplàncton costaner més gran, com les diatomees.[49][52][53][54] La gran resposta de la floració i el canvi de la comunitat han donat lloc a preocupacions ambientals sobre la fertilització de grans seccions de regions HNLC. Un estudi suggereix que les diatomees creixen preferentment durant els experiments de fertilització. Algunes diatomees, com la pseudo-nitzschia, alliberen la neurotoxina àcid domoic, enverinant els peixos que pasturen.[53] Si les diatomees creixen preferentment durant els experiments de fertilització de ferro, les fertilitzacions sostingudes podrien augmentar la intoxicació per àcid domoic a la xarxa alimentària marina propera a les zones fertilitzades.[53]
La pols eòlica
modifica
El ferro entra a les regions HNLC remotes a través de dos mètodes principals: l'afegiment d'aigua rica en nutrients i la deposició de pols atmosfèrica. El ferro ha de ser reposat amb freqüència i en formes biodisponibles a causa de la seva insolubilitat, ràpida captació a través de sistemes biològics i afinitat d'enllaç amb lligands.[55][56] És possible que la deposició de pols no resulti en floracions de fitoplàncton, tret que la pols de sedimentació estigui en la forma de ferro biodisponible correcta. A més, s'ha de dipositar ferro durant les estacions productives i coincidir amb les relacions RKR adequades dels nutrients superficials.[19][57] La pols eòlica té una influència més gran en les regions HNLC de l'hemisferi nord perquè hi ha més massa de terra que contribueix a la deposició de pols.[58] A causa de l'aïllament de l'oceà Antàrtic de la terra, l'enfonsament relacionat amb la difusió turbulenta proporciona ferro a les regions HNLC.[59]
Hipòtesi de control del pasturatge
modificaFormulat per John Walsh el 1976, la hipòtesi de pasturatge assenyala que el pasturatge per heteròtrofs suprimeix la productivitat primària en àrees d'alta concentració de nutrients.[46][60] La depredació per microzooplancton representa principalment la pèrdua de fitoplàncton a les regions HNLC. El pasturatge mitjançant un zooplàncton més gran i la barreja advectiva també són responsables d'una petita proporció de pèrdues en les comunitats de fitoplàncton.[6][61][62] El pasturatge constant limita el fitoplàncton a un estoc constant. Sense aquesta pressió del pasturatge, alguns científics creuen que el fitoplàncton petit produiria floracions malgrat l'esgotament de micronutrients, ja que el fitoplàncton més petit sol tenir requeriments inferiors de ferro i pot absorbir nutrients a un ritme més lent.[55][61]
Punt de vista contemporani
modificaEl consens científic actual acorda que les àrees HNLC no tenen una alta productivitat a causa d'una combinació de ferro i limitacions fisiològiques, la pressió del pasturatge i forçaments físics.[2][6][48][54][61][63] El grau en què cada factor contribueix a la baixa producció pot diferir en cada regió HNLC. La limitació del ferro permet que el fitoplàncton més petit i més ferro-frugal creixi a ritmes ràpids, mentre que el pasturatge per microzooplàncton manté estocs estables d'aquests fitoplàncton més petits.[6][56][61] Un cop disponibles els micronutrients, el pasturatge pot limitar les mides de la floració.[46][48][49][51][54] Les limitacions addicionals de micronutrients dels oligoelements, com el zinc o el cobalt, poden suprimir la floració del fitoplàncton.[12] La barreja turbulenta a les regions HNLC de latitud alta (Pacífic nord i oceà Antàrtic) pot barrejar el fitoplàncton per sota de la profunditat crítica necessària per tenir un creixement comunitari.[46]
Geoenginyeria a les regions HNLC
modificaTeoria
modifica
Atès que els experiments de fertilització amb ferro passats han donat lloc a grans floracions de fitoplàncton, alguns han suggerit que s'han de dur a terme experiments de fertilització oceànica a gran escala per treure el diòxid de carboni antropogènic inorgànic en forma de partícules de carboni orgànic. La fertilització estimularia la productivitat biològica, provocant una disminució de la quantitat de diòxid de carboni superficial inorgànic dins d'una zona fertilitzada. La floració moriria i suposadament s'enfonsaria a l'oceà profund, portant gran part del carboni absorbit al fons marí i segrestant-lo del cicle de carboni a curt termini en l'oceà profund o als sediments oceànics.[51][64][65][66][67]
Eficiència i eficàcia
modificaPer eliminar eficaçment el carboni antropogènic de l'atmosfera, la fertilització amb ferro hauria de provocar una eliminació significativa de les partícules de carboni de l'oceà superficial i transportar-lo a l'oceà profund.[47][48][65][66] Diversos estudis han estimar que menys del 7-10% del carboni absorbit durant una floració seria segrestat,[68] i que una fertilització global amb ferro sostinguda només disminuiria 15-25 ppm del diòxid de carboni atmosfèric.[7][65] La quantitat de diòxid de carboni eliminat pot ser compensada pel cost de combustible per adquirir, transportar i alliberar quantitats significatives de ferro a les regions HNLC remotes.[66]
Existeixen moltes preocupacions mediambientals per a la fertilització amb ferro a gran escala. Mentre que les floracions poden ser estudiades i traçades, els científics encara no saben si la producció addicional s'incorporaria a la cadena alimentària o cauria al fons de l'oceà després que es produeixi una floració.[47][48] Fins i tot, si s'exporta carboni a la profunditat pot crear zones anòxiques o provocar una acidificació de les aigües profundes.[66][69] S'han observat canvis pronunciats de la comunitat a les diatomees durant la fertilització, i encara no està clar si el canvi en la composició d'espècies té efectes ambientals a llarg termini.[53][66]
Referències
modifica- ↑ 1,0 1,1 1,2 Lalli, C.M; Parsons, T.R. Biological Oceanography: An Introduction (en anglès). 2. Burlington, MA: Elsevier Butterworth Heinemann, 2004, p. 55.
- ↑ 2,0 2,1 2,2 Pitchford, J.W.; Brindley, J. «Iron limitation, grazing pressure and oceanic high-nutrient-low chlorophyll (HNLC) regions» (en anglès). publicació of Plankton Research, 21(3), 1999, pàg. 525–547. DOI: 10.1093/plankt/21.3.525.
- ↑ 3,0 3,1 Venables, H; Moore, C. M «Phytoplankton and light limitation in the Southern Ocean: Learning from high-nutrient, high-chlorophyll areas» (en anglès). J. Geophys. Res., 115, C02015, 2010. DOI: 10.1029/2009JC005361.
- ↑ 4,0 4,1 Hassler, C. S.; Sinoir, M.; Clementson, L. A.; Butler, E. C. V. «Exploring the Link between Micronutrients and Phytoplankton in the Southern Ocean during the 2007 Austral Summer» (en anglès). Frontiers in Microbiology, 3, 2012, pàg. 202. DOI: 10.3389/fmicb.2012.00202. PMC: 3392650. PMID: 22787456.
- ↑ 5,0 5,1 5,2 Martin, John. Primary productivity and biogeochemical cycles in the sea (en anglès). Springer US, 1992, p. 122–137.
- ↑ 6,00 6,01 6,02 6,03 6,04 6,05 6,06 6,07 6,08 6,09 6,10 Martin, John; Gordon, Michael; Fitzwater, Steve; Broenkow, William W. «VERTEX: phytoplankton/iron studies in the Gulf of Alaska» (en anglès). Deep Sea Research Part A. Oceanographic Research Papers, 35(6=, 1989, pàg. 649–680.
- ↑ 7,0 7,1 7,2 7,3 Miller, Charles B.; Wheeler, Patricia A. Biological oceanography (en anglès). Chichester, West Sussex: John Wiley and Sons, Ltd., 2012, p. 49–62. ISBN 9781444333015. OCLC 794619582.
- ↑ Redfield, A.C.; Ketchum, G.H.; Richards, F.A.. «The influence of organisms on the composition of sea water». A: The Sea (en anglès). Nova York: Wiley-Interscience, 1963, p. 26–77.
- ↑ Werner, Stumm; Morgan, James J. Aquatic Chemistry : Chemical Equilibria and Rates in Natural Waters (en anglès). 3. Hoboken: Wiley, 2013. ISBN 9780471673033. OCLC 863203908.
- ↑ 10,0 10,1 «Iron and zinc effects on silicic acid and nitrate uptake kinetics in three high-nutrient, low-chlorophyll (HNLC) regions» (
 PDF) (en anglès). ResearchGate.
PDF) (en anglès). ResearchGate.
- ↑ 11,0 11,1 11,2 Anderson, M. A.; Morel, F. M. M.; Guillard, R. R. L. «Growth limitation of a coastal diatom by low zinc ion activity» (en anglès). Nature, 276(5683), 2 novembre 1978-11, pàg. 70–71. Bibcode: 1978Natur.276...70A. DOI: 10.1038/276070a0.
- ↑ 12,0 12,1 12,2 De La Rocha, Christina L.; Hutchins, David A.; Brzezinski, Mark A.; Zhang, Yaohong «Effects of iron and zinc deficiency on elemental composition and silica production by diatoms» (en anglès). Marine Ecology Progress Series, 195, 2000, pàg. 71–79. Bibcode: 2000MEPS..195...71D. DOI: 10.3354/meps195071. JSTOR: 24855011.
- ↑ Dugdale, Richard C.; Wilkerson, Frances P.; Minas, Hans J. «The role of a silicate pump in driving new production» (en anglès). Deep Sea Research Part I: Oceanographic Research Papers, 42(5), 1r maig 1995, pàg. 697–719. Bibcode: 1995DSRI...42..697D. DOI: 10.1016/0967-0637(95)00015-X.
- ↑ 14,0 14,1 14,2 14,3 Dugdale, Richard C.; Wilkerson, Frances P. «Silicate regulation of new production in the equatorial Pacific upwelling» (en anglès). Nature, 391(6664), 15-01-1998, pàg. 270–273. Bibcode: 1998Natur.391..270D. DOI: 10.1038/34630.
- ↑ Cullen, John J.; Lewis, Marlon R.; Davis, Curtiss O.; Barber, Richard T. «Photosynthetic characteristics and estimated growth rates indicate grazing is the proximate control of primary production in the equatorial Pacific» (en anglès). Geophysical Research: Oceans, 9(C1), 15-01-1992, pàg. 639–654. Bibcode: 1992JGR....97..639C. DOI: 10.1029/91JC01320. ISSN: 2156-2202.
- ↑ Landry, M. R.; Constantinou, J.; Latasa, M.; Brown, S. L.; et al. «Biological response to iron fertilization in the eastern equatorial Pacific (IronEx II). III. Dynamics of phytoplankton growth and microzooplankton grazing» (en anglès). Marine Ecology Progress Series, 201, 09-08-2000, pàg. 57–72. Bibcode: 2000MEPS..201...57L. DOI: 10.3354/meps201057.
- ↑ Oeste, Franz Dietrich; Richter, Renaud de; Ming, Tingzhen; Caillol, Sylvain «Climate engineering by mimicking natural dust climate control. The iron salt aerosol method» (en anglès). Earth Syst. Dynam., 8 (1), S., 2017, pàg. 1–54. DOI: 10.5194/esd-8-1-2017.
- ↑ 18,0 18,1 18,2 Miller, Charles B.; Wheeler, Patricia A. «Wiley: Biological Oceanography» (en anglès). www.wiley.com.
- ↑ 19,0 19,1 Boyd, P.W.; Mackie, D.S.; Hunter, K.A. «Aerosol iron deposition to the surface ocean — Modes of iron supply and biological responses» (en anglès). Marine Chemistry, 120(1–4), 2010, pàg. 128–143. DOI: 10.1016/j.marchem.2009.01.008.
- ↑ Hansard, S. P.; Landing, W. M.; Measures, C. I.; Voelker, B. M. «Dissolved iron (II) in the Pacific Ocean: measurements from the PO2 and P16N CLIVAR/CO 2 repeat hydrography expeditions» (en anglès). Deep Sea Research Part I: Oceanographic Research Papers, 56(7), 2009, pàg. 1117–1129. Bibcode: 2009DSRI...56.1117H. DOI: 10.1016/j.dsr.2009.03.006.
- ↑ Lam, Phoebe J.; Bishop, James K. B. «The continental margin is a key source of iron to the HNLC North Pacific Ocean» (en anglès). Geophysical Research Letters, 35(7), 1r abril 2008, pàg. L07608. Bibcode: 2008GeoRL..35.7608L. DOI: 10.1029/2008gl033294. ISSN: 1944-8007.
- ↑ 22,0 22,1 Tyrrell, T.; Merico, A.; Waniek, J. J.; Wong, C. S.; et al. «Effect of seafloor depth on phytoplankton blooms in high-nitrate, low-chlorophyll (HNLC) regions» (en anglès). publicació of Geophysical Research: Biogeosciences, 110(G2), 1r desembre 2005, pàg. G02007. Bibcode: 2005JGRG..110.2007T. DOI: 10.1029/2005jg000041. ISSN: 2156-2202.
- ↑ 23,0 23,1 Langmann, B.; Zakšek, K.; Hort, M.; Duggen, S. «Volcanic ash as fertiliser for the surface ocean» (en anglès). Atmospheric Chemistry and Physics, 10(8), 2010, pàg. 3891–3899. DOI: 10.5194/acp-10-3891-2010.
- ↑ 24,0 24,1 Pondaven, P.; Ruiz-Pino, D.; Druon, J.N.; Fravalo, C.; Tréguer, P. «Factors controlling silicon and nitrogen biogeochemical cycles in high nutrient, low chlorophyll systems (the Southern Ocean and the North Pacific): Comparison with a mesotrophic system (the North Atlantic)» (en castellà). Deep Sea Research Part I: Oceanographic Research Papers, 46(11), 1999, pàg. 1923–1968. Bibcode: 1999DSRI...46.1923P. DOI: 10.1016/s0967-0637(99)00033-3.
- ↑ Chavez, F P.; Toggweiler, J R. «Physical estimates of global new production: The upwelling contribution». A: 'Dahlem Workshop on Upwelling in the Ocean: Modern Processes and Ancient Records (en anglès). Chichester, UK: John Wiley & Sons, 1995, p. 313-320.
- ↑ Eppley, R. W.; Renger, E. H. «Nitrate utilization by plankton in the equatorial Pacific March 1988 along 150°W» (en angès). publicació of Geophysical Research: Oceans, 97(C1), 15-01-1992, pàg. 663–668. Bibcode: 1992JGR....97..663E. DOI: 10.1029/91JC01271. ISSN: 2156-2202.
- ↑ 27,0 27,1 27,2 27,3 27,4 Winckler, Gisela; Anderson, Robert F.; Jaccard, Samuel L.; Marcantonio, Franco «Ocean dynamics, not dust, have controlled equatorial Pacific productivity over the past 500,000 years» (en anglès). Proceedings of the National Academy of Sciences, 113(22), 31-05-2016, pàg. 6119–6124. Bibcode: 2016PNAS..113.6119W. DOI: 10.1073/pnas.1600616113. ISSN: 0027-8424. PMC: 4896667. PMID: 27185933.
- ↑ 28,0 28,1 28,2 Landry, Michael R.; Selph, Karen E.; Taylor, Andrew G.; Décima, Moira; et al. «Phytoplankton growth, grazing and production balances in the HNLC equatorial Pacific» (en anglès). Deep Sea Research Part II: Topical Studies in Oceanography, 58(3), 1r febrer 2011, pàg. 524–535. Bibcode: 2011DSRII..58..524L. DOI: 10.1016/j.dsr2.2010.08.011.
- ↑ Chisholm, F. M.; Morel. What controls phytoplankton production in nutrient-rich areas of the open sea? (en anglès), 1991.
- ↑ Pollard, Raymond; Tréguer, Paul; Read, Jane «Quantifying nutrient supply to the Southern Ocean» (en anglès). publicació of Geophysical Research, 111(C5), 2006, pàg. C05011. Bibcode: 2006JGRC..111.5011P. DOI: 10.1029/2005JC003076.
- ↑ Morrison, Adele K.; Frölicher, Thomas L.; Sarmiento, Jorge L. «Upwelling in the Southern Ocean» (en anglès). Physics Today, 68(1), 2015, pàg. 27–32. Bibcode: 2015PhT....68a..27M. DOI: 10.1063/PT.3.2654.
- ↑ Sañudo-Wilhelmy, S.; Flegal, A. R. «Potential influence of Saharan dust on the chemical composition of the Southern Ocean» (en anglès). Geochem. Geophys. Geosyst., 4, 2003, pàg. 1063, doi:10.1029/2003GC000507, 7.
- ↑ de Baar, Hein J. W.; Boyd, Philip W.; Coale, Kenneth H.; Landry, Michael R.; et al. «Synthesis of iron fertilization experiments: From the Iron Age in the Age of Enlightenment» (en anglès). publicació of Geophysical Research: Oceans, 110(C9), 1r setembre 2005, pàg. C09S16. Bibcode: 2005JGRC..110.9S16D. DOI: 10.1029/2004jc002601. ISSN: 2156-2202.
- ↑ Martin, John H.; Gordon, R. Michael; Fitzwater, Steve E. «Iron in Antarctic waters» (en anglès). Nature, 345(6271), 10-05-1990, pàg. 156–158. Bibcode: 1990Natur.345..156M. DOI: 10.1038/345156a0. ISSN: 1476-4687.
- ↑ Pollard, R.; Sanders, R.; Lucas, M.; Statham, P. «The Crozet natural iron bloom and export experiment (CROZEX)» (en anglès). Deep-Sea Res., II: Top. Stud. Oceanogr. 54, 1905–1914, 2007, pàg. 18-20.
- ↑ Blain, S.; Quéguiner, B.; Trull, T.W. «The natural iron fertilization experiment KEOPS (KErguelen Ocean and Plateau compared Study)» (en anglès). Deep-Sea Res. II 55, 2008, pàg. 559–565.
- ↑ 37,0 37,1 37,2 Charette, Matt; Sanders, Richard; Zhou, Meng «Southern Ocean natural iron fertilization» (en angles). Deep-Sea Research II, 90, 2011, pàg. 283.
- ↑ Venables, Hugh; Moore, C. Mark «Phytoplankton and light limitation in the Southern Ocean: Learning from high-nutrient, high-chlorophyll areas» (en anglès). publicació of Geophysical Research: Oceans, 115(C2), 1r febrer 2010, pàg. C02015. Bibcode: 2010JGRC..115.2015V. DOI: 10.1029/2009JC005361. ISSN: 2156-2202.
- ↑ 39,0 39,1 Charette, Matt; Sanders, Richard; Zhou, Meng «Southern Ocean natural iron fertilization» ( PDF) (en anglès). WHOI, 90, 2011, pàg. 283.
- ↑ 40,0 40,1 Liggett, Daniela; Storey, Bryan; Cook, Yvonne; Meduna, Veronika. Exploring the cognom continent: an introduction to Antarctica (en anglès). Springer, 2015.
- ↑ Hart, T. J.. On the phytoplankton of the south-west Atlantic and the Bellingshausen Sea, 1929-31 (en anglès). VIII, 1934, p. 1-268.
- ↑ Weier, John «John Martin (1935-1993)» (en anglès). On the Shoulders of Giants.
- ↑ Boyd,, P.W.; et al. «Mesoscale iron enrichment experiments 1993-2005: synthesis and future directions.» (en anglès). Science, 2007, pàg. 315.
- ↑ Martin, P.; van der Loeff, M.R.; Cassar, N.; Vandromme, P.; d'Ovidio, F.; Stemmann, L.; Rengarajan, R.; Soares, M.; González, H.E.; Ebersbach, F.; Lampitt, R.S.; Sanders, R.; Barnett, B.A.; Smetacek, V.; Naqvi, S.W.A «Iron fertilization enhanced net community production but not downward particle flux during the Southern Ocean iron fertilization experiment LOHAFEX» (en anglès). Global Biogeochemical Cycles, 27(3), 2013, pàg. 871-881. DOI: 10.1002/gbc.20077. ISSN: 1944-9224.
- ↑ «Ocean-fertilization project off Canada sparks furore» (en anglès). Nature News & Comment.
- ↑ 46,0 46,1 46,2 46,3 Martin, J.H.; Coale, K.H.; Johnson, K.S.; Fitzwater, S.E.; et al. «Testing the iron hypothesis in ecosystem of the equatorial Pacific Ocean» (en anglès). Nature, 371(6493), 9-1994, pàg. 123–129. Bibcode: 1994Natur.371..123M. DOI: 10.1038/371123a0.
- ↑ 47,0 47,1 47,2 47,3 47,4 Fujii, Masahiko; Yoshie, Naoki; Yamanaka, Yasuhiro; Chai, Fei «Simulated biogeochemical responses to iron enrichments in three high nutrient, low chlorophyll (HNLC) regions» (en anglès). Progress in Oceanography, 64(2-4), 2005, pàg. 307–324. Bibcode: 2005PrOce..64..307F. DOI: 10.1016/j.pocean.2005.02.017.
- ↑ 48,0 48,1 48,2 48,3 48,4 Edwards, Andrew M.; Platt, Trevor; Sathyendranath, Shubha «The high-nutrient, low-chlorophyll regime of the ocean: limits on biomass and nitrate before and after iron enrichment» (en anglès). Ecological Modelling, 171(1-2), 2004, pàg. 103–125. DOI: 10.1016/j.ecolmodel.2003.06.001.
- ↑ 49,0 49,1 49,2 49,3 Behrenfeld, Michael J. «Confirmation of iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean» (en anglès). Nature, 383(6600), 10-10-1996, pàg. 508–511. Bibcode: 1996Natur.383..508B. DOI: 10.1038/383508a0.
- ↑ Martin, John H.; Fitzwater, Steve E.; Gordon, R. Michael «Iron deficiency limits phytoplankton growth in Antarctic waters» (en anglès). Global Biogeochemical Cycles, 4(1), 1r març 1990, pàg. 5-12. Bibcode: 1990GBioC...4....5M. DOI: 10.1029/gb004i001p00005. ISSN: 1944-9224.
- ↑ 51,0 51,1 51,2 «IPCC - Intergovernmental Panel on Climate Change» (en anglès). www.ipcc.ch.
- ↑ Lam, Phoebe J.; Tortell, Philippe D.; Morel, Francois M.M. «Differential effects of iron additions on organic and inorganic carbon production by phytoplankton» (en anglès). Limnology and Oceanography, 46(5), 3-2001, pàg. 1199-1202. Bibcode: 2001LimOc..46.1199L. DOI: 10.4319/lo.2001.46.5.1199.
- ↑ 53,0 53,1 53,2 53,3 Trick, Charles G.; Bill, Brian D.; Cochlan, William P.; Wells, Mark L.; et al. «Iron enrichment stimulates toxic diatom production in high-nitrate, low-chlorophyll areas» (en anglès). Proceedings of the National Academy of Sciences, 107(13), 30-03-2010, pàg. 5887-5892. Bibcode: 2010PNAS..107.5887T. DOI: 10.1073/pnas.0910579107. ISSN: 0027-8424. PMC: 2851856. PMID: 20231473.
- ↑ 54,0 54,1 54,2 Tsuda, Atsushi; Takeda; Saito; Nishioka «Evidence for the grazing hypothesis: grazing reduces phytoplankton responses of the HNLC ecosystem to iron enrichment in the Western Subarctic Pacific (SEEDS II)» (en anglès). publicació of Oceanography, 63(6), 7-2007, pàg. 983–994. DOI: 10.1007/s10872-007-0082-x.
- ↑ 55,0 55,1 Mioni, Cécile E.; Handy, Sara M.; Ellwood, Michael J.; Twiss, Michael R.; et al. «Tracking changes in bioavailable Fe within high-nitrate low-chlorophyll oceanic waters: A nom estimate using a heterotrophic bacterial bioreporter» (en anglès). Global Biogeochemical Cycles, 19(4), 1r desembre 2005, pàg. GB4S25. Bibcode: 2005GBioC..19.4S25M. DOI: 10.1029/2005gb002476. ISSN: 1944-9224.
- ↑ 56,0 56,1 Pitchford, Jonathan William; Brindley, John «Iron limitation, grazing pressure and oceanic high nutrient-low chlorophyll (HNLC) regions» (en anglès). Plankton Research, 21(3), 1999, pàg. 525-547. DOI: 10.1093/plankt/21.3.525.
- ↑ Jickells, T. D.; An, Z. S.; Andersen, K. K.; Baker, A. R.; et al. «Global Iron Connections Between Desert Dust, Ocean Biogeochemistry, and Climate» (en anglès). Science, 308(5718), 1r abbril 2005, pàg. 67-71. Bibcode: 2005Sci...308...67J. DOI: 10.1126/science.1105959. ISSN: 0036-8075. PMID: 15802595.
- ↑ Behrenfeld, Michael J.; Kolber, Zbigniew S. «Widespread iron limitation of phytoplankton in the South Pacific Ocean» (en anglès). Science, 283(5403), 05-02-1999, pàg. 840-843. Bibcode: 1999Sci...283..840B. DOI: 10.1126/science.283.5403.840. PMID: 9933166.
- ↑ Wagener, Thibaut; Guieu, Cécile; Losno, Rémi; Bonnet, Sophie; Mahowald, Natalie «Revisiting atmospheric dust export to the Southern Hemisphere ocean: Biogeochemical implications». Global Biogeochemical Cycles, 22(2), 1r juny 2008, pàg. GB2006. Bibcode: 2008GBioC..22.2006W. DOI: 10.1029/2007gb002984. ISSN: 1944-9224.
- ↑ Walsh, John J. «Herbivory as a factor in patterns of nutrient utilization in the sea» (en anglès). Limnology and Oceanography, 21(1), 1r gener 1976, pàg. 1-13. Bibcode: 1976LimOc..21....1W. DOI: 10.4319/lo.1976.21.1.0001. ISSN: 1939-5590.
- ↑ 61,0 61,1 61,2 61,3 Frost, Bruce W. «The role of grazing in nutrient-rich areas of the open sea» (en anglès). Limnology and Oceanography, 38(8), 1991, pàg. 1616-1630. Bibcode: 1991LimOc..36.1616F. DOI: 10.4319/lo.1991.36.8.1616.
- ↑ Minas, Hans Joachim; Minas, Monique «Net community production in "High Nutrient-Low Chlorophyll" waters of the tropical and Antarctic Oceans: grazing vs iron hypothesis» (en anglès). Oceanologica Acta, 15(2), 1992, pàg. 145-162.
- ↑ Frost, B.W. «Grazing control of phytoplankton stock in the open subarctic Pacific Ocean: a model assessing the role of mesozooplankton, particularly the large calanoid copepods Neocalanus spp.» (en anglès). Marine Ecology Progress Series, 39, 27-07-1987, pàg. 49–68. Bibcode: 1987MEPS...39...49F. DOI: 10.3354/meps039049.
- ↑ Behrenfeld, Michael J.; Bale, Anthony J.; Kolber, Zbigniew S.; Aiken, James; Falkowski, Paul G. «Confirmation of iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean» (en anglès). Nature, 383(6600), 10-1996, pàg. 508-511. Bibcode: 1996Natur.383..508B. DOI: 10.1038/383508a0.
- ↑ 65,0 65,1 65,2 Zeebe, R. E.; Archer, D. «Feasibility of ocean fertilization and its impact on future atmospheric CO2 levels» (en anglès). Geophysical Research Letters, 32(9), 1r maig 2005, pàg. L09703. Bibcode: 2005GeoRL..32.9703Z. DOI: 10.1029/2005gl022449. ISSN: 1944-8007.
- ↑ 66,0 66,1 66,2 66,3 66,4 «Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, 2007» (en anglès). www.ipcc.ch. Arxivat de l'original el 2014-10-12. [Consulta: 25 juliol 2019].
- ↑ Waller, Rhian «Iron Fertilization: Savoir to Climate Change or Ocean Dumping?» (en anglès). Geographic, 18-10-2012.
- ↑ Boyd, Philip W.; Law, Cliff S.; Wong, C.S.; Nojiri, Yukihiro; et al. «The decline and fate of an iron-induced subarctic phytoplankton bloom» (en anglès). Nature, 428(6982), 17-03-2004, pàg. 549-553. Bibcode: 2004Natur.428..549B. DOI: 10.1038/nature02437. ISSN: 1476-4687. PMID: 15058302.
- ↑ Cao, Long; Caldeira, Ken «Can ocean iron fertilization mitigate ocean acidification?» (en anglès). Climatic Change, 99(1-2), 1r març 2010, pàg. 303–311. DOI: 10.1007/s10584-010-9799-4. ISSN: 0165-0009.